NAD+: A Research Overview
A research-library overview of nicotinamide adenine dinucleotide (NAD+): its dinucleotide chemistry, dual redox and signaling roles, salvage and de novo biosynthesis, subcellular compartmentalization, and a discovery history spanning 1906 to the sirtuin era. Educational reference.

Introduction
Nicotinamide adenine dinucleotide (NAD+) is a pyridine dinucleotide coenzyme present in every living cell. Unlike most compounds catalogued in the research-peptide literature, NAD+ is not a peptide, a hormone, or a receptor ligand; it is a small-molecule cofactor whose defining feature is that it participates in two mechanistically unrelated classes of reaction. In one, it shuttles reducing equivalents through central metabolism without being consumed. In the other, it is cleaved and spent as a substrate by a family of signaling enzymes. This overview summarizes NAD+ chemistry, its redox and signaling duality, its biosynthetic routes, its subcellular organization, and the historical record of its discovery, in a neutral educational register.
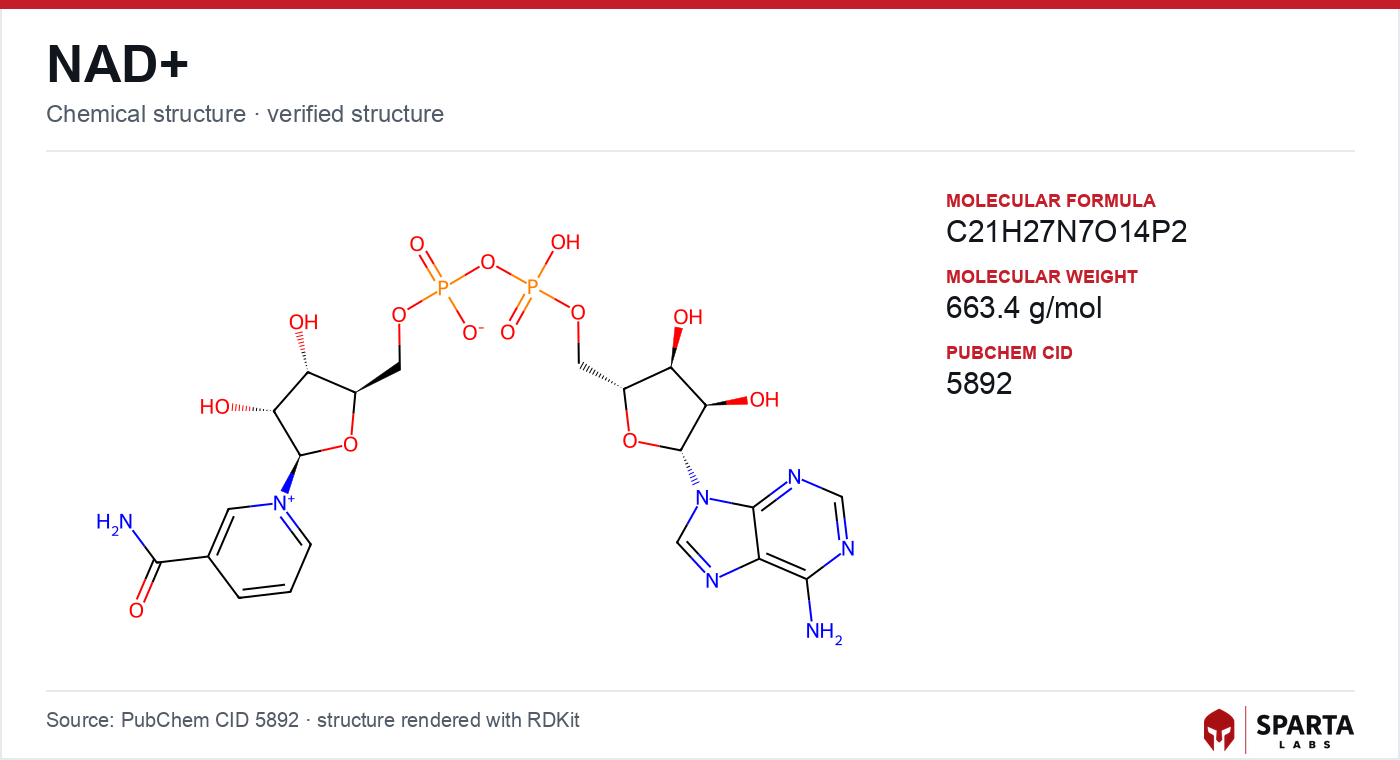
Figure: chemical structure of NAD+.
Chemistry and Dinucleotide Structure
NAD+ (CAS 53-84-9; molecular formula C₂₁H₂₇N₇O₁₄P₂; molecular weight approximately 663.4 g/mol) is a dinucleotide built from two nucleoside monophosphate units joined by a 5',5'-pyrophosphate bridge. One half is adenosine 5'-monophosphate (AMP); the other is nicotinamide mononucleotide (NMN), in which the nicotinamide base is attached to ribose. The pyrophosphate linkage and the two ribose sugars form the backbone, while the adenine and nicotinamide rings project from opposite ends of the molecule.
The nicotinamide ring is the chemically active center. Its pyridinium nitrogen makes the ring electron-poor, allowing the C4 position to accept a hydride ion (H⁻) and form the reduced species, NADH. This two-electron, one-hydride transfer is reversible, which is why the NAD+/NADH couple can operate as a recyclable electron carrier. At physiological pH the molecule carries net negative charge from its phosphate groups and does not readily cross intact lipid membranes.
A closely related molecule, NADP+, carries an additional phosphate at the 2' position of the adenosine ribose. That single modification reroutes it into a largely separate enzymatic network associated with reductive biosynthesis and antioxidant regeneration. Because NAD+ and NADP+ serve distinct pools of enzymes despite their structural similarity, this article restricts its scope to the NAD+/NADH couple.
Two Roles: Redox Cofactor Versus Consumed Substrate
The single most distinctive property of NAD+ is that it operates through two chemically separate mechanisms. Distinguishing them is essential to reading the literature accurately.
In its redox role, NAD+ accepts hydride from metabolic intermediates during catabolism, forming NADH, and is later regenerated when NADH is reoxidized, for example at the mitochondrial electron transport chain. In this cycle NAD+ is not net-consumed; the same molecules turn over repeatedly. This is the classical textbook function that places NAD+ at the center of glycolysis, the citric acid cycle, and oxidative phosphorylation.
In its signaling role, a separate set of enzymes cleaves the glycosidic bond between nicotinamide and ribose, spending the NAD+ molecule as a substrate rather than recycling it. Reviews by Verdin and by Cantó, Menzies, and Auwerx catalogued this consumption by three enzyme families: the sirtuins (NAD+-dependent deacylases), the poly(ADP-ribose) polymerases (PARPs, associated with DNA-damage responses), and the cyclic ADP-ribose synthases such as CD38 [1,2]. Each reaction releases free nicotinamide and draws down the total cellular NAD+ pool, which is why sustained signaling activity creates ongoing demand for NAD+ resynthesis.
Findings from research models do not establish safety or efficacy in humans. Sparta Labs makes no claims about the use of this compound.
The enzymatic reading of NAD+ concentration is what links a metabolic cofactor to gene regulation and the DNA-damage response. Related signaling pathways in mitochondrial biology are examined in the MOTS-c research overview, a mitochondria-derived peptide studied in the same metabolic context.
Biosynthesis: Salvage, De Novo, and Preiss-Handler Routes
Because signaling enzymes continuously consume NAD+, cells maintain three convergent biosynthetic routes rather than relying on a static pool.
The salvage pathway recycles the nicotinamide released by sirtuins, PARPs, and CD38 back into NAD+. Nicotinamide phosphoribosyltransferase (NAMPT) converts nicotinamide to NMN, and NMN adenylyltransferases (NMNATs) then join NMN to AMP to regenerate NAD+. In most mammalian tissues this salvage loop, rather than fresh synthesis, accounts for the bulk of NAD+ turnover.
The de novo pathway builds the nicotinamide ring from the amino acid tryptophan through the kynurenine pathway, ultimately feeding into NAD+ synthesis. The Preiss-Handler pathway, characterized biochemically by Preiss and Handler in 1958, describes the route from dietary nicotinic acid (niacin) through nicotinic acid mononucleotide and nicotinic acid adenine dinucleotide to NAD+ [3].
Because intact NAD+ is poorly taken up by cells, research on manipulating the intracellular NAD+ pool has focused on precursors that enter these pathways more readily, including nicotinamide riboside (NR) and NMN. In a clinical pharmacokinetic study, Trammell and colleagues reported that oral NR administration produced measurable, dose-related elevations in the human blood NAD+ metabolome, providing early human data on precursor bioavailability [4].
Subcellular Compartmentalization
NAD+ is not distributed uniformly through the cell. It is partitioned into distinct pools in the cytosol, the mitochondria, and the nucleus, each served by relatively separate enzymatic machinery and each maintained by its own NMNAT isoform. The mitochondrial pool tends to be the largest in energetically demanding tissues such as cardiac and skeletal muscle, consistent with the organelle's role in oxidative phosphorylation.
This compartmentalization matters for interpreting research: an intervention that changes NAD+ in one pool does not necessarily change it in another, and whole-cell or whole-blood measurements can obscure pool-specific dynamics. The recognition that the nuclear pool is directly coupled to sirtuin and PARP activity helped connect NAD+ availability to chromatin regulation and the DNA-damage response, and it remains an active area of biochemical investigation.
Pharmacological Classification and Regulatory Status
In biochemical and pharmacological literature NAD+ is classified as a dinucleotide coenzyme and pyridine nucleotide. It is an endogenous molecule present in all mammalian tissues and does not hold FDA approval as a therapeutic drug for any indication, consistent with its handling as a research compound.
The surrounding precursor landscape has its own regulatory dimension. NR is marketed in the United States as a dietary-supplement ingredient under an FDA New Dietary Ingredient acknowledgment. NMN's status has been the subject of ongoing industry and regulatory dialogue following FDA actions that treated it differently from established supplement ingredients. These distinctions apply to specific precursor molecules and their intended commercial uses, not to NAD+ as a research reagent. Research-grade NAD+ from Sparta Labs is supplied with a batch-specific certificate of analysis; verification standards for this material are described in the NAD+ sourcing and quality reference.
Discovery History
The record of NAD+ research spans more than a century. In 1906 Arthur Harden and William John Young identified a heat-stable, dialysable cofactor in yeast extract, which they termed "cozymase," as a requirement for fermentation [5]. Harden shared the 1929 Nobel Prize in Chemistry with Hans von Euler-Chelpin, whose work helped establish that cozymase was a dinucleotide. In the 1930s Otto Heinrich Warburg identified the nicotinamide ring as the hydride-accepting moiety, cementing NAD+'s place at the center of respiratory metabolism.
The biosynthetic routes from dietary niacin were mapped by Preiss and Handler in 1958 [3]. A distinct research era opened in 2000, when Imai, Armstrong, Kaeberlein, and Guarente reported in Nature that the yeast silencing protein Sir2 is an NAD+-dependent histone deacetylase, directly tying NAD+ availability to chromatin regulation and gene expression [6]. That finding reframed NAD+ from a purely metabolic cofactor into a signaling molecule and helped launch the sirtuin field. A fuller chronology appears in the NAD+ discovery and research history article, and the enzymatic mechanisms are detailed in the NAD+ mechanism of action reference.
References
-
Verdin E. NAD+ in aging, metabolism, and neurodegeneration. Science. 2015;350(6265):1208–1213. DOI: 10.1126/science.aac4854. Link
-
Cantó C, Menzies KJ, Auwerx J. NAD+ metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metab. 2015;22(1):31–53. PMC4487780. Link
-
Preiss J, Handler P. Biosynthesis of diphosphopyridine nucleotide. I. Identification of intermediates. J Biol Chem. 1958;233(2):488–492. Link
-
Trammell SA, Schmidt MS, Weidemann BJ, Redpath P, Jaksch F, Dellinger RW, et al. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016;7:12948. DOI: 10.1038/ncomms12948. Link
-
Harden A, Young WJ. The alcoholic ferment of yeast-juice. Proc R Soc Lond B. 1906;77(519):405–420. DOI: 10.1098/rspb.1906.0029. Link
-
Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795–800. DOI: 10.1038/35001622. Link
Frequently asked questions
What is NAD+ and what makes it unusual among research compounds?
Nicotinamide adenine dinucleotide (NAD+) is a pyridine dinucleotide coenzyme found in every living cell. It is unusual because it operates through two mechanistically separate roles: as a recyclable hydride carrier in redox metabolism, and as a consumed substrate cleaved by signaling enzymes such as sirtuins and PARPs. It is a small-molecule cofactor, not a peptide, hormone, or receptor ligand.
What is the chemical structure of NAD+?
NAD+ is a dinucleotide made of two nucleoside monophosphate units, adenosine 5'-monophosphate and nicotinamide mononucleotide, joined by a 5',5'-pyrophosphate bridge. Its molecular formula is C21H27N7O14P2 with a molecular weight near 663.4 g/mol (CAS 53-84-9). The nicotinamide ring is the reactive center, accepting a hydride at C4 to form NADH.
How does NAD+ differ from NADP+?
NADP+ carries an extra phosphate group at the 2' position of the adenosine ribose that NAD+ lacks. Although the two molecules are structurally similar, this single difference routes them into largely separate enzymatic networks. NAD+ predominates in catabolic redox reactions and as a substrate for sirtuins and PARPs, while NADP+ is associated with reductive biosynthesis.
How is NAD+ synthesized in cells?
Cells maintain NAD+ through three convergent routes. The salvage pathway recycles nicotinamide released by NAD+-consuming enzymes via NAMPT and NMNAT enzymes. The de novo pathway builds the nicotinamide ring from tryptophan through the kynurenine pathway, and the Preiss-Handler pathway derives NAD+ from dietary nicotinic acid. Salvage accounts for most NAD+ turnover in mammalian tissues.
Is NAD+ approved by the FDA?
NAD+ is an endogenous coenzyme and does not hold FDA approval as a therapeutic drug for any indication, consistent with its handling as a research compound. Separately, the precursor nicotinamide riboside is marketed as a dietary-supplement ingredient under an FDA New Dietary Ingredient acknowledgment, while nicotinamide mononucleotide's regulatory status has been the subject of ongoing dialogue.