NAD+: Discovery and Research History
Tracing NAD+ across four scientific eras: the 1906 discovery of cozymase in yeast fermentation, the 1930s convergence of redox chemistry and niacin nutrition, the Preiss-Handler biosynthetic pathway, and the sirtuin and precursor research that followed. Educational reference.

For research use only. Not for human consumption. This article is educational reference material. It is not medical advice and is not a recommendation to use any substance.
Introduction
Few molecules in biochemistry have been studied continuously for as long as nicotinamide adenine dinucleotide (NAD+). Its documented research record extends back to 1906 and passes through four scientific communities that arrived at the same coenzyme from very different starting points: fermentation chemists, nutrition scientists studying pellagra, enzymologists mapping cellular respiration, and, most recently, molecular biologists studying chromatin and aging. Each community redefined what NAD+ was understood to be, and each redefinition changed the questions researchers asked of it. This article traces that lineage chronologically, with attention to the primary papers that marked each transition. A chemistry and regulatory summary is covered in the accompanying NAD+ research overview.
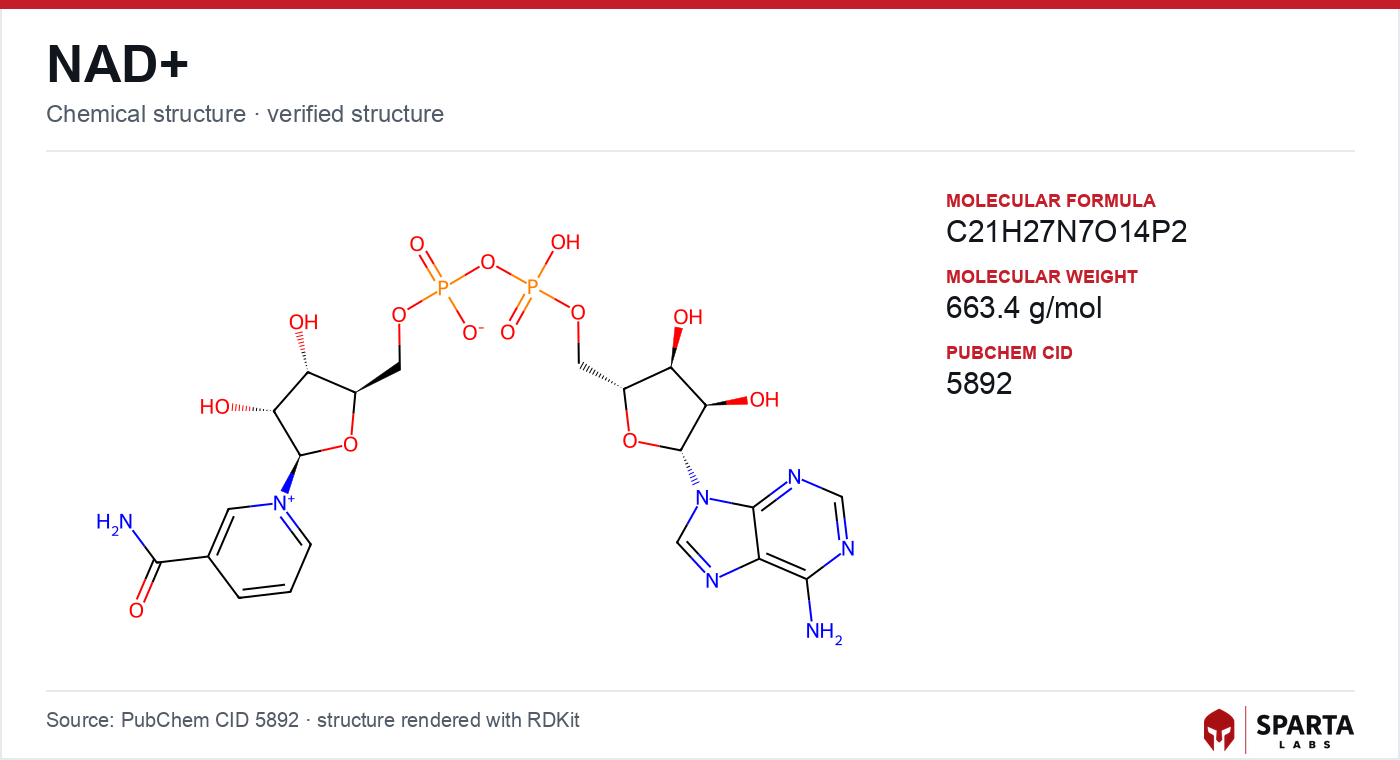
Figure: chemical structure of NAD+.
Cozymase: A Fermentation Cofactor Without a Structure (1906–1929)
NAD+ entered the scientific record before its structure, its name, or its function were known. Arthur Harden and William John Young, working at the Lister Institute in London, were studying alcoholic fermentation in cell-free yeast extracts when they encountered an anomaly. In a 1906 report to the Proceedings of the Royal Society of London, they described that adding boiled yeast extract to fresh yeast juice accelerated fermentation, even though boiling destroyed the enzymatic proteins the extract contained [1]. The implication was that fermentation required not only heat-labile proteins but also a heat-stable component of a different chemical nature.
A companion paper the same year established that this stable factor was dialysable: it passed through a semipermeable membrane that retained proteins, marking it as a small molecule rather than a protein [2]. Harden and Young called the factor "cozymase" and defined it as a required cofactor for yeast fermentation. The term captured a functional role rather than a chemical identity, which remained undetermined for more than two decades.
Hans von Euler-Chelpin subsequently devoted years to characterizing cozymase, establishing that it was a nucleotide-containing molecule built from two nucleotide units joined through a phosphate bridge — a dinucleotide. In 1929, Harden and von Euler-Chelpin shared the Nobel Prize in Chemistry "for their investigations on the fermentation of sugar and fermentative enzymes," the first formal recognition of the molecule the world would later call NAD+.
Two 1930s Discoveries: Redox Chemistry and the Niacin Link
The 1930s resolved cozymase into a defined chemical entity along two independent research fronts that converged on the same molecule.
The first front was mechanistic. Otto Heinrich Warburg, already a Nobel laureate for his work on cellular respiration, turned to the pyridine coenzymes. His group established that the nicotinamide ring of the coenzyme was the chemically active moiety, and that it accepted a hydride ion at the C4 position of the pyridine ring during biological oxidation reactions. This identified NAD+ as a hydride carrier and placed it at the mechanistic center of cellular energy metabolism, a role it retains in every modern biochemistry curriculum. The closely related phosphorylated coenzyme NADP+ was recognized in the same period, defining a family of pyridine nucleotide cofactors partitioned between catabolic and biosynthetic redox chemistry.
The second front was nutritional. Pellagra — a deficiency disease then epidemic across the American South and marked by dermatitis, diarrhea, and dementia — had resisted explanation. Conrad Elvehjem and colleagues at the University of Wisconsin reported in 1937 that nicotinic acid resolved "blacktongue," a canine disease analogous to human pellagra. This connected the redox coenzyme to dietary biology: nicotinic acid (niacin) and its amide nicotinamide were dietary precursors of NAD+, and niacin was thereby defined as vitamin B3, a compound whose dietary absence produces a defined disease syndrome. The molecule studied by fermentation chemists and the vitamin sought by nutrition scientists were the same.
Mapping Biosynthesis: The Preiss–Handler Pathway (1950s)
Once NAD+ was recognized as a vitamin-derived coenzyme, the enzymatic route from dietary niacin to the finished dinucleotide became a defined research problem. Jack Preiss and Philip Handler addressed it in two 1958 papers in the Journal of Biological Chemistry, mapping the multi-step conversion of nicotinic acid to NAD+ through intermediates including nicotinic acid mononucleotide and nicotinic acid adenine dinucleotide [3,4]. The route they described became known as the Preiss–Handler pathway and remained the canonical account of NAD+ biosynthesis from dietary niacin for decades. Their work fixed the biosynthetic framework against which later-discovered routes — the tryptophan-derived de novo pathway and, much later, the nicotinamide riboside route — would be positioned as alternatives.
From Cofactor to Consumed Substrate: ADP-Ribosylation and Sirtuins (1960s–2000)
Through the middle of the twentieth century NAD+ was understood almost entirely as a recyclable redox cofactor, reduced to NADH and reoxidized without net consumption. That picture changed when researchers found enzymes that cleave NAD+ and use its ADP-ribose moiety as a building block, consuming the coenzyme in the process. In the 1960s and 1970s, work on poly(ADP-ribose) polymerase (PARP) established that ADP-ribose units are transferred from NAD+ onto acceptor proteins in reactions linked to DNA-damage signaling. For the first time, NAD+ was a substrate that could be depleted, not merely a carrier that was regenerated — implying that its cellular availability could rate-limit signaling pathways.
The most consequential reframing came in 2000, when Imai, Armstrong, Kaeberlein, and Guarente reported in Nature that Sir2, a yeast silencing protein tied to replicative lifespan, was an NAD-dependent histone deacetylase [5].
Findings from research models do not establish safety or efficacy in humans. Sparta Labs makes no claims about the use of this compound.
That single paper repositioned NAD+ from a metabolic bookkeeping molecule to a regulatory signal that couples cellular energy status to chromatin structure and gene regulation. Because the sirtuin family is evolutionarily conserved — from bacteria through the human SIRT1–SIRT7 enzymes — the finding was immediately relevant to mammalian cell biology and became one of the most-cited results in the field. The receptor-level and enzymatic detail of this coupling is examined in the NAD+ mechanism of action article. The same era characterized CD38, previously studied in immunology, as a major cellular NADase. The recognition that PARPs, sirtuins, and CD38 all draw on a shared, finite NAD+ pool gave the coenzyme a role at the intersection of DNA repair, metabolic sensing, and immune regulation.
The Precursor Era: Nicotinamide Riboside and Clinical Pharmacology (2004–Present)
A distinct research phase opened when Bieganowski and Brenner reported in Cell in 2004 that nicotinamide riboside (NR) is a dietary NAD+ precursor taken up through a specific kinase-mediated route, the NRK pathway, that bypasses both the Preiss–Handler and de novo pathways [6]. NR could therefore raise cellular NAD+ independently of nicotinic acid or nicotinamide — a distinction with direct implications for the precursor pharmacology that followed.
The precursor concept met the sirtuin-and-aging literature to produce a wave of preclinical work. Cantó and colleagues reported in 2012 that dietary NR was associated with altered mitochondrial biology and sirtuin activity in a mouse model [7]. Gomes, Sinclair, and colleagues described in 2013 a decline in tissue NAD+ during aging in mice linked to disrupted nuclear-mitochondrial communication, with partial reversal reported after precursor administration in that model [8]. These animal findings motivated translation into human pharmacokinetics.
The first human characterization of oral NR came from Trammell, Schmidt, and colleagues in 2016, who documented dose-dependent increases in blood NAD+ metabolites following oral administration in a first-in-human study [9]. Subsequent controlled trials extended this work across healthy and older adults, establishing short-term tolerability and confirming that NR and related precursors measurably shift the blood NAD+ metabolome in humans. The functional metabolic outcomes reported across those trials are summarized in the accompanying NAD+ published research article.
The Current Research Landscape
By the mid-2020s, NAD+ research spans mechanistic biology and human clinical investigation. A methodological milestone has been non-invasive measurement of NAD+ in living human tissue by magnetic resonance spectroscopy, which allows researchers to track the coenzyme in the brain without biopsy — a capability that opens neurological research applications previously inaccessible. Active clinical registrations touch cardiovascular, hepatic, and neurodegenerative research questions, while mechanistic work continues to ask which NAD+-consuming system most governs tissue NAD+ dynamics: PARP activation from accumulated DNA damage, CD38 upregulation in aged tissue, or salvage-pathway flux through NAMPT.
The scientific arc of NAD+ is also visible in adjacent mitochondrial biology. The mitochondria-derived peptide MOTS-c follows a comparably recent characterization timeline, and the antioxidant tripeptide glutathione shares NAD+'s history of being isolated from cellular extracts long before its regulatory roles were understood. NAD+ from Sparta Labs is offered as research-grade material with batch documentation. As tissue-level measurement methods mature and precursor trials accumulate, NAD+ remains a field defined more by open questions than settled answers.
References
-
Harden A, Young WJ. The alcoholic ferment of yeast-juice. Proc R Soc Lond B. 1906;77(519):405–420. DOI: 10.1098/rspb.1906.0029. Link
-
Harden A, Young WJ. The alcoholic ferment of yeast-juice. Part II — The coferment of yeast-juice. Proc R Soc Lond B. 1906;78(526):369–375. DOI: 10.1098/rspb.1906.0070. Link
-
Preiss J, Handler P. Biosynthesis of diphosphopyridine nucleotide. I. Identification of intermediates. J Biol Chem. 1958;233(2):488–492. PMID: 13563526. Link
-
Preiss J, Handler P. Biosynthesis of diphosphopyridine nucleotide. II. Enzymatic aspects. J Biol Chem. 1958;233(2):493–500. PMID: 13563527. Link
-
Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795–800. DOI: 10.1038/35001622. PMID: 10693811. Link
-
Bieganowski P, Brenner C. Discoveries of nicotinamide riboside as a nutrient and conserved NRK genes establish a Preiss-Handler independent route to NAD+ in fungi and humans. Cell. 2004;117(4):495–502. DOI: 10.1016/S0092-8674(04)00416-7. PMID: 15137942. Link
-
Cantó C, Houtkooper RH, Pirinen E, Youn DY, Oosterveer MH, Cen Y, et al. The NAD+ precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012;15(6):838–847. DOI: 10.1016/j.cmet.2012.04.022. PMID: 22682224. Link
-
Gomes AP, Price NL, Ling AJ, Moslehi JJ, Montgomery MK, Rajman L, et al. Declining NAD+ induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013;155(7):1624–1638. DOI: 10.1016/j.cell.2013.11.037. PMID: 24360282. Link
-
Trammell SA, Schmidt MS, Weidemann BJ, Redpath P, Jaksch F, Dellinger RW, et al. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016;7:12948. DOI: 10.1038/ncomms12948. PMID: 27721479. Link
Disclaimer. Statements in this article have not been evaluated by the Food and Drug Administration. This compound is not intended to diagnose, treat, cure, or prevent any disease. Sparta Labs sells research-use-only materials. Content is provided for educational and informational purposes only and does not constitute medical advice. Consult a qualified medical professional for any health concerns.
Frequently asked questions
When was NAD+ first discovered?
NAD+ was first observed in 1906, when Arthur Harden and William John Young at the Lister Institute in London identified a heat-stable, dialysable cofactor in yeast extract required for fermentation, which they named cozymase. Its dinucleotide structure was later characterized by Hans von Euler-Chelpin, and Harden and von Euler-Chelpin shared the 1929 Nobel Prize in Chemistry for the work.
How is NAD+ connected to niacin and pellagra?
In 1937, Conrad Elvehjem and colleagues at the University of Wisconsin reported that nicotinic acid resolved blacktongue, a canine disease analogous to human pellagra. This established niacin (vitamin B3) and its amide nicotinamide as dietary precursors of NAD+, linking the fermentation coenzyme to nutritional biology.
What is the Preiss-Handler pathway?
The Preiss-Handler pathway is the enzymatic route by which dietary nicotinic acid is converted into NAD+. Jack Preiss and Philip Handler mapped it in two 1958 papers in the Journal of Biological Chemistry, describing intermediates including nicotinic acid mononucleotide and nicotinic acid adenine dinucleotide. It remained the canonical framework for NAD+ biosynthesis for decades.
Why did the 2000 Imai et al. Nature paper matter for NAD+ research?
Imai, Armstrong, Kaeberlein, and Guarente reported in 2000 that the yeast silencing protein Sir2 was an NAD-dependent histone deacetylase. This repositioned NAD+ from a recyclable redox cofactor to a regulatory signal linking cellular energy status to chromatin and gene regulation, and it launched a large body of sirtuin research because the enzyme family is conserved from bacteria to humans.
What is nicotinamide riboside (NR) and when was it identified as an NAD+ precursor?
Nicotinamide riboside is a dietary NAD+ precursor. Bieganowski and Brenner reported in Cell in 2004 that NR is taken up through a distinct kinase-mediated route, the NRK pathway, that bypasses the Preiss-Handler and de novo pathways, allowing cellular NAD+ to be raised independently of nicotinic acid or nicotinamide.