Glutathione: Published Research Summary
A structured review of peer-reviewed glutathione research, organized by the questions each body of work set out to answer: biosynthesis and pool size, intracellular compartmentalization, GPx enzymatic chemistry, S-glutathionylation signaling, and controlled oral-supplementation trials.

For research use only. Not for human consumption. This article is educational reference material. It is not medical advice and is not a recommendation to use any substance.
Introduction
Glutathione (GSH; γ-L-glutamyl-L-cysteinyl-glycine) is a low-molecular-weight thiol tripeptide that has served as a reference system for redox biochemistry for more than a century. Its unusual gamma-glutamyl peptide bond, its millimolar intracellular abundance, and the reactivity of its single cysteine thiol have made it a recurring subject across biochemistry, cell biology, and human nutrition research. This article summarizes representative peer-reviewed studies on glutathione, organized by the questions each body of work set out to answer rather than by a fixed template. It reports what investigators published, with attribution and methodology context, and does not draw independent conclusions. The reported molecular chemistry underlying these findings is treated separately in the glutathione mechanism of action article.
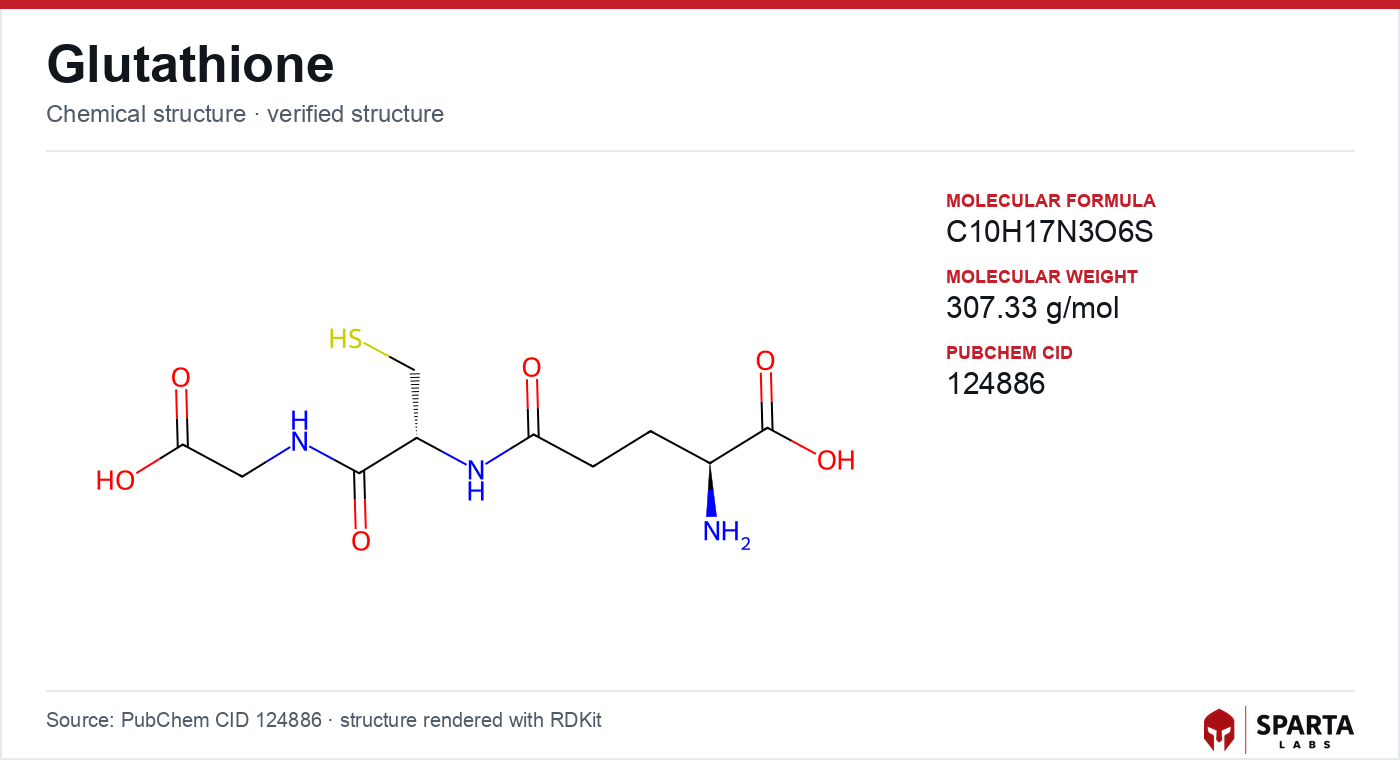
Figure: chemical structure of glutathione (γ-L-glutamyl-L-cysteinyl-glycine).
How the Glutathione Literature Is Organized
Because glutathione touches many subfields, the published record is best understood as several parallel research programs that rarely use the same methods. Foundational biochemistry work characterized GSH biosynthesis and the enzymes that consume it, using purified proteins and kinetic assays. Cell-biology work asked where GSH actually sits inside a cell and how its redox state is regulated, relying on fluorescent probes and mass-spectrometry proteomics. A separate signaling literature examined reversible protein modification by glutathione. Finally, a clinical-nutrition literature tested whether ingested glutathione registers as a measurable change in body pools. The sections below trace each of these programs through landmark reviews and primary trials.
Foundational Biochemistry: Biosynthesis and Pool Size
The framing review for modern glutathione biochemistry is that of Forman, Zhang, and Rinna, published in Molecular Aspects of Medicine in 2009 [1]. Synthesizing several decades of enzymology, the authors reported that intracellular GSH concentrations range from roughly 0.5 to 10 mM across cell types, with hepatocytes at the upper end. They described the two-step, ATP-dependent biosynthetic pathway catalyzed by glutamate-cysteine ligase (GCL) and glutathione synthetase (GS), identified GCL as the rate-limiting step, and summarized evidence that cysteine availability and GCL activity are the principal determinants of intracellular content. The review also noted that plasma GSH concentrations are orders of magnitude lower than intracellular levels, a consequence of limited export and rapid extracellular turnover.
Findings from research models do not establish safety or efficacy in humans. Sparta Labs makes no claims about the use of this compound.
This biosynthetic framing matters for interpreting later work: because GSH is made intracellularly from constituent amino acids rather than absorbed intact in large quantities, the relationship between ingested glutathione and tissue glutathione has been a central and non-trivial question, addressed in the clinical section below.
Compartmentalization: Where Glutathione Actually Sits
For decades the intracellular GSH pool was modeled as a single well-mixed reservoir. Baty, Hampton, and Winterbourn challenged that simplification in a 2014 Redox Biology study using fluorescent glutathione probes [2]. The investigators reported that intracellular glutathione is heterogeneously distributed, with total glutathione in the endoplasmic reticulum exceeding roughly 15 mM under the conditions studied while the integrated whole-cell concentration measured near 7 mM. They described a glutathione concentration gradient across the ER membrane. This work supported a compartment-specific view of cellular redox regulation in which the oxidizing environment of the secretory pathway and the reducing environment of the cytosol are maintained as distinct redox domains.
The compartmentalization theme was extended in a 2014 Frontiers in Pharmacology review by Aquilano, Baldelli, and Ciriolo [3]. Drawing on cell-based experiments, the authors reported that cytoplasmic, mitochondrial, and nuclear glutathione pools appear to be regulated independently and may serve distinct roles. They discussed proposals that nuclear GSH participates in chromatin dynamics and protection of DNA against oxidative strand breaks, and characterized the glutathione and thioredoxin systems as two independent but partially overlapping NADPH-dependent redox networks. These reviews reframed glutathione from a bulk antioxidant buffer to a spatially organized regulatory system, a shift that continues to inform live-cell sensor studies.
Enzymatic Chemistry: Glutathione Peroxidases
Much of glutathione's reported antioxidant function is enzymatically mediated, and the selenoenzyme glutathione peroxidase-1 (GPx-1) is the most-studied representative. Lubos, Loscalzo, and Handy reviewed GPx-1 in Antioxidants & Redox Signaling in 2011 [4]. The authors characterized GPx-1 as an intracellular selenoenzyme that reduces hydrogen peroxide and soluble lipid hydroperoxides using GSH as the electron donor, with the active-site selenocysteine cycling through selenol, selenenic acid, and mixed-diselenide intermediates. They reviewed selenium-dependent translational regulation of GPx-1 expression and summarized human genetic-association data linking GPx-1 activity to cardiovascular and metabolic outcomes, while explicitly noting that associations do not establish causality.
A distinct GPx isoform, GPx4, has become central to a newer research area. GPx4 is selective for lipid hydroperoxides embedded within membranes and uses GSH as its reducing substrate. Published research characterized GPx4 as a regulator of ferroptosis, a form of iron-dependent regulated cell death, with loss of GPx4 activity reported to trigger ferroptotic death in cell models. This connection has made the glutathione–GPx4 axis a focus of contemporary cancer-biology and cell-death research, and it illustrates how a classical antioxidant co-substrate acquired an unexpected role in a modern signaling field.
Reversible Signaling: Protein S-Glutathionylation
Beyond serving as an enzyme co-substrate, glutathione forms reversible mixed disulfides with protein cysteine residues, a modification termed S-glutathionylation. Xiong, Uys, Tew, and Townsend reviewed this chemistry comprehensively in Antioxidants & Redox Signaling in 2011 [5]. The authors described the documented routes of formation: spontaneous thiol-disulfide exchange with oxidized glutathione (GSSG), reaction of GSH with sulfenic-acid intermediates generated by partial cysteine oxidation, and glutaredoxin-catalyzed pathways. They catalogued a large set of proteins reported to undergo S-glutathionylation in cell-based experiments, including transcription factors, kinases, metabolic enzymes, and structural proteins, and framed the modification as a candidate mechanism for oxidative-stress-responsive regulation of specific signaling nodes.
For most proteins in the published S-glutathionylation catalogs, the functional consequence of the modification on activity, localization, or interaction has not yet been experimentally resolved. That gap between inventory and function is one of the more tractable open problems in the field and continues to generate proteomic and biochemical follow-on studies.
Clinical Nutrition: Does Ingested Glutathione Register in Body Pools
The most translational strand of the literature asks a deceptively simple question: does oral glutathione change measurable glutathione in the body, given that biosynthesis is intracellular and gastrointestinal hydrolysis is expected. Richie and colleagues addressed this in a randomized controlled trial published in the European Journal of Nutrition in 2015 [6]. The trial enrolled 54 healthy adults who received oral GSH at two levels or placebo over six months. The authors reported statistically significant differences in glutathione concentrations measured in erythrocytes, plasma, and lymphocytes relative to baseline and placebo at various timepoints, along with differences in buccal-cell glutathione. The trial provided early controlled evidence that oral glutathione formulations can be associated with measurable changes in peripheral glutathione parameters.
Sinha and colleagues extended this line with a pilot trial of liposomal oral glutathione in 12 healthy adults, published in the European Journal of Clinical Nutrition in 2018 [7]. The liposomal formulation was designed to address the hydrolysis limitations of standard oral glutathione. The authors reported statistically significant changes in whole-blood and erythrocyte glutathione relative to baseline, along with changes in natural-killer-cell cytotoxicity as one measured parameter, while explicitly noting that the small sample size and short duration limited generalizability. Together these trials frame formulation and bioavailability as the practical variables that distinguish glutathione from constituent amino acids as an ingested compound.
Knowledge Gaps and the Active Research Frontier
The published record leaves several questions open. The bioavailability of different glutathione formulations remains an active comparison, with the Richie and Sinha trials serving as early data points that larger and longer studies are intended to build upon. The physiological significance of compartment-specific glutathione pools in humans, as opposed to the cell lines and animal models where most mechanistic data originate, is being addressed with genetically encoded live-cell glutathione sensors. And the functional annotation of S-glutathionylation targets remains largely incomplete.
Glutathione also sits within a broader cellular-metabolism research cluster, where redox balance intersects with mitochondrial energetics and cofactor biology. Readers surveying that cluster may find the NAD+ research overview relevant to the NADPH-dependent recycling that regenerates reduced glutathione, and the MOTS-c research overview relevant to mitochondria-centered metabolic signaling. For background on glutathione's classification and discovery timeline, the glutathione research overview and the glutathione discovery history provide complementary context. Sparta Labs stocks glutathione for laboratory research applications, characterized to the analytical standards described elsewhere in this library.
References
-
Forman HJ, Zhang H, Rinna A. Glutathione: overview of its protective roles, measurement, and biosynthesis. Mol Aspects Med. 2009;30(1-2):1-12. PMID: 18796312. DOI: 10.1016/j.mam.2008.08.006. PubMed
-
Baty JW, Hampton MB, Winterbourn CC. Intracellular glutathione pools are heterogeneously concentrated. Redox Biol. 2013;1(1):508-513. PMID: 24251119. DOI: 10.1016/j.redox.2013.10.005. PubMed
-
Aquilano K, Baldelli S, Ciriolo MR. Glutathione: new roles in redox signaling for an old antioxidant. Front Pharmacol. 2014;5:196. PMID: 25206336. DOI: 10.3389/fphar.2014.00196. PubMed
-
Lubos E, Loscalzo J, Handy DE. Glutathione peroxidase-1 in health and disease: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2011;15(7):1957-1997. PMID: 21087145. DOI: 10.1089/ars.2010.3586. PubMed
-
Xiong Y, Uys JD, Tew KD, Townsend DM. S-glutathionylation: from molecular mechanisms to health outcomes. Antioxid Redox Signal. 2011;15(1):233-270. PMID: 20919933. DOI: 10.1089/ars.2010.3540. PubMed
-
Richie JP Jr, Nichenametla S, Neidig W, Calcagnotto A, Haley JS, Schell TD, et al. Randomized controlled trial of oral glutathione supplementation on body stores of glutathione. Eur J Nutr. 2015;54(2):251-263. PMID: 24791752. DOI: 10.1007/s00394-014-0706-z. PubMed
-
Sinha R, Sinha I, Calcagnotto A, Trushin N, Haley JS, Schell TD, et al. Oral supplementation with liposomal glutathione elevates body stores of glutathione and markers of immune function. Eur J Clin Nutr. 2018;72(1):105-111. PMID: 28853742. DOI: 10.1038/ejcn.2017.132. PubMed
Disclaimer. Statements in this article have not been evaluated by the Food and Drug Administration. This compound is not intended to diagnose, treat, cure, or prevent any disease. Sparta Labs sells research-use-only materials. Content is provided for educational and informational purposes only and does not constitute medical advice. Consult a qualified medical professional for any health concerns.
Frequently asked questions
How is glutathione made inside cells?
Published biochemistry reviews describe a two-step, ATP-dependent biosynthetic pathway catalyzed by glutamate-cysteine ligase (GCL) and glutathione synthetase (GS). GCL is characterized as the rate-limiting step, and cysteine availability together with GCL activity are reported as the principal determinants of intracellular glutathione content. Intracellular concentrations range from roughly 0.5 to 10 mM across cell types.
Is glutathione distributed evenly inside a cell?
Research using fluorescent glutathione probes reported that intracellular glutathione is heterogeneously distributed rather than uniformly mixed. Baty and colleagues observed endoplasmic-reticulum glutathione exceeding roughly 15 mM under their experimental conditions while whole-cell concentration measured near 7 mM. Separate reviews describe cytoplasmic, mitochondrial, and nuclear pools as independently regulated.
What did controlled trials of oral glutathione report?
Richie and colleagues conducted a six-month randomized controlled trial in 54 healthy adults and reported statistically significant differences in glutathione measured in erythrocytes, plasma, and lymphocytes relative to baseline and placebo. A later pilot trial by Sinha and colleagues in 12 adults reported changes in whole-blood and erythrocyte glutathione with a liposomal formulation, while noting sample-size limitations.
What is S-glutathionylation?
S-glutathionylation is a reversible modification in which glutathione forms a mixed disulfide with a protein cysteine residue. Published reviews describe several documented formation routes and catalog many proteins reported to undergo the modification in cell-based experiments. It has been framed as a candidate mechanism for oxidative-stress-responsive regulation of specific signaling proteins.
How does glutathione relate to ferroptosis research?
The glutathione peroxidase isoform GPx4 is selective for membrane-embedded lipid hydroperoxides and uses glutathione as its reducing substrate. Published research characterized GPx4 as a regulator of ferroptosis, an iron-dependent form of regulated cell death, with loss of GPx4 activity reported to trigger ferroptotic death in cell models. This axis is a focus of contemporary cell-death and cancer-biology research.