Melanotan-2: Mechanism of Action
A mechanism-focused reference on Melanotan-2 (MT-II): its lactam-bridged cyclic scaffold, four-subtype melanocortin receptor engagement, Gs-cAMP transduction, and the central-versus-peripheral signaling questions that remain open in the literature. Educational reference.

For research use only. Not for human consumption. This article is educational reference material. It is not medical advice and is not a recommendation to use any substance.
Introduction
Melanotan-2 (MT-II) is a synthetic, cyclic analog of the endogenous melanocortin peptide alpha-melanocyte-stimulating hormone (alpha-MSH). It was engineered as a conformationally constrained, superpotent melanotropin whose pharmacology is defined by non-selective agonism across most of the melanocortin receptor family. Understanding its reported mechanism therefore means tracing a single ligand as it engages several distinct G-protein-coupled receptors, each expressed in different tissues and coupled to overlapping but not identical intracellular outputs. This article summarizes the reported molecular pharmacology of MT-II drawn from peer-reviewed primary literature. Background on its chemistry, nomenclature, and pharmacological classification is covered in the Melanotan-2 research overview, and a broader survey of the preclinical study record appears in the Melanotan-2 published research summary.
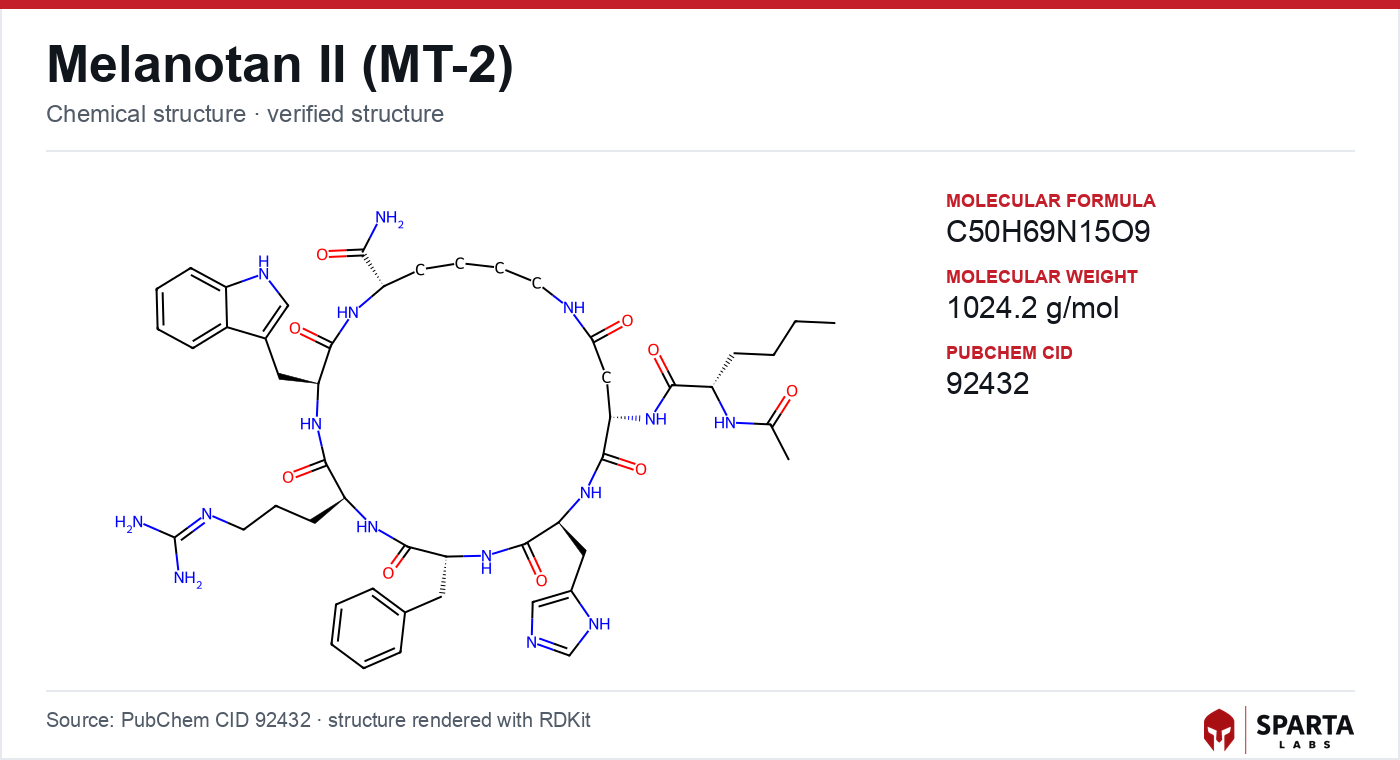
Figure: chemical structure of Melanotan II.
The Cyclic Scaffold and Its Pharmacophore
The mechanistic starting point for MT-II is its architecture. Native alpha-MSH is a linear tridecapeptide, and its melanocortin activity depends on a compact core motif, the sequence His-Phe-Arg-Trp corresponding to positions 6 through 9 of the alpha-MSH chain [1]. Structure-activity work on melanotropins identified this tetrapeptide as the minimal message sequence recognized by melanocortin receptors, so any effort to build a more potent, more stable agonist had to preserve and stabilize that core [2].
MT-II accomplishes this through two design choices. First, a D-phenylalanine substitution replaces the native L-Phe at the message-sequence position, a stereochemical inversion associated in melanotropin chemistry with increased receptor potency and resistance to peptidase inactivation [2]. Second, an intramolecular lactam bridge cyclizes the backbone, locking the His-D-Phe-Arg-Trp pharmacophore into a rigid, receptor-competent conformation rather than allowing it to sample the flexible ensemble a linear peptide adopts in solution. Reviews of constrained melanocortin ligands describe this conformational restriction as a general strategy for converting a flexible, rapidly degraded hormone into a long-lived, high-affinity probe [2].
The practical consequence is that MT-II presents a pre-organized pharmacophore to the receptor. Because entropy loss on binding is reduced when the ligand is already shaped correctly, the constrained scaffold is associated with higher affinity than the linear parent peptide, a relationship documented across the melanotropin structure-activity literature [2].
Engagement Across Four Receptor Subtypes
The melanocortin system comprises five G-protein-coupled receptors, MC1R through MC5R, each encoded by a distinct gene and each showing a characteristic tissue distribution [1]. MT-II is reported as a non-selective agonist across four of these subtypes: MC1R, MC3R, MC4R, and MC5R. This breadth is the defining feature of its mechanism and the reason preclinical observations attributed to MT-II span such different physiological domains, from pigment biology to central circuitry [3].
The one subtype MT-II is not reported to meaningfully engage is MC2R. MC2R is the adrenocorticotropic hormone (ACTH) receptor, and its recognition requires N-terminal residues present in the full 39-residue ACTH molecule but absent from the compact melanocortin message sequence [1]. Because MT-II is built around only the core pharmacophore, it lacks the structural determinants MC2R demands, and this exclusion is mechanistically consistent with the receptor's known ligand requirements.
Non-selectivity is a double-edged property in mechanistic interpretation. In an intact organism, a systemic exposure to MT-II activates every accessible subtype at once, so any observed physiological outcome reflects the summed, sometimes competing, contributions of multiple receptors in multiple tissues. Researchers therefore rely on subtype-selective antagonists and genetic knockout models to assign specific outputs to specific receptors, a methodological point revisited below.
Gs-cAMP Transduction: The Shared Second-Messenger Route
Across its target subtypes, MT-II is reported to activate the canonical Gs-coupled signaling cascade shared by the melanocortin receptors [1][3]. Agonist binding stabilizes the active receptor conformation, which catalyzes guanine-nucleotide exchange on the Galpha-s subunit. The activated Galpha-s dissociates from the Gbeta-gamma dimer and stimulates adenylyl cyclase, raising intracellular cyclic adenosine monophosphate (cAMP). Elevated cAMP then activates protein kinase A (PKA), and in appropriate cell types PKA phosphorylates the transcription factor CREB, linking receptor occupancy to changes in gene expression.
A 2019 review in Molecules by Sharma and colleagues examined MC4R activation in detail and described Gs-cAMP signaling as the principal transduction route through which MC4R agonists exert their documented preclinical pharmacology [4]. The same review noted that MC4R is not a purely Gs-coupled receptor: it can also recruit ERK1/2 mitogen-activated protein kinase signaling, and the relationship between the cAMP and MAPK arms remains an area of continuing mechanistic work.
Findings from research models do not establish safety or efficacy in humans. Sparta Labs makes no claims about the use of this compound.
Because the second-messenger route is shared, the tissue in which a receptor sits, rather than the signaling machinery itself, largely determines the physiological readout. The following two sections illustrate this by contrasting the two most-studied subtypes for MT-II, one predominantly peripheral and one predominantly central.
MC1R and the Pigment-Cell cAMP Axis
MC1R is expressed prominently on melanocytes, the pigment-producing cells of the skin, and it is the receptor most directly connected to melanocortin biology's original discovery in pigmentation research [1]. In melanocytes, Gs-cAMP signaling downstream of MC1R engages the microphthalmia-associated transcription factor (MITF) program that governs the melanin-synthetic machinery, a pathway central to how these cells respond to melanocortin agonism [1][5].
MT-II has served in this setting as a stable pharmacological probe of MC1R-driven signaling in melanocyte-lineage systems. Because its cyclic scaffold resists the rapid degradation that limits native alpha-MSH, it provides a durable agonist for characterizing the MC1R-cAMP-MITF axis in vitro and in preclinical models [2][5]. The mechanistic point of interest is that the same Gs-cAMP cascade described above, when it operates in a pigment cell rather than a hypothalamic neuron, produces a transcriptional rather than a neurophysiological output, underscoring how receptor localization dictates the biological meaning of an otherwise conserved signal.
MC4R and Central Melanocortin Circuitry
MC4R presents the contrasting case. It is expressed heavily in the central nervous system, particularly within hypothalamic nuclei that participate in energy-balance regulation, and it is one of the most intensively studied melanocortin receptors in neuroscience [1][4]. The Sharma 2019 review situates MC4R at the center of the central melanocortin system, where its Gs-cAMP signaling contributes to the circuitry that integrates energy-state information [4].
Foundational work on the melanocortin system by Cone and by Mountjoy established that POMC-derived peptides and their receptors form a regulatory node in the hypothalamus, with MC4R functioning as a key relay whose disruption alters central signaling in animal models [3][6]. Because MT-II is a non-selective agonist that reaches this circuitry, it has been employed as a research tool to probe MC4R-dependent central signaling. Attributing a given central observation specifically to MC4R, however, requires selective antagonists or receptor-null models, since MC3R is also present in overlapping regions and MT-II activates both [3][4].
This central-versus-peripheral contrast is the clearest illustration of MT-II's mechanistic character: a single, structurally simple ligand distributes across anatomically separated receptor populations, and the physiology that emerges depends on which population dominates in a given experimental preparation. A structurally related melanocortin ligand developed for a narrower receptor profile, PT-141 (Bremelanotide), illustrates how modifying the same scaffold shifts the subtype-engagement pattern, and the alpha-MSH-derived fragment KPV represents a further structurally distinct member of the same peptide family whose signaling has been studied in complementary contexts.
Open Mechanistic Questions
Several questions remain unresolved in the MT-II and broader melanocortin literature. The most consequential follows directly from non-selectivity: because MT-II simultaneously engages MC1R, MC3R, MC4R, and MC5R in intact systems, cleanly partitioning any systemic outcome among its receptor targets continues to depend on selective antagonists, knockout models, and subtype-selective comparator ligands.
A second question concerns signaling bias. Whether MT-II preferentially couples a given receptor to the Gs-cAMP arm versus beta-arrestin or MAPK routes, and whether that preference varies by subtype or tissue, is an active topic in G-protein-coupled receptor pharmacology, and the ERK1/2 coupling of MC4R noted by Sharma and colleagues is one facet of it [4]. A third concerns species translation: much of the mechanistic data derives from rodent models, and how faithfully that pharmacology maps onto primate receptor biology remains under study [3][4]. Researchers evaluating MT-II as a characterization tool can review third-party analytical standards for the material through the Melanotan-2 product listing.
References
-
Gantz I, Fong TM. The melanocortin system. Am J Physiol Endocrinol Metab. 2003;284(3):E468-74. PMID: 12556347. DOI: 10.1152/ajpendo.00524.2002. PubMed
-
Hruby VJ. Design of cyclic peptides with biological activities from biologically active peptides: the case of peptide modulators of melanocortin receptors. Biopolymers. 2016;106(6):884-8. PMID: 27486849. DOI: 10.1002/bip.22950. PubMed
-
Cone RD. Anatomy and regulation of the central melanocortin system. Nat Neurosci. 2005;8(5):571-8. PMID: 15856065. DOI: 10.1038/nn1455. PubMed
-
Sharma S, Garfield AS, Shah B, Kleyn P, Numao S, Costello DA, et al. Current mechanistic and pharmacodynamic understanding of melanocortin-4 receptor activation. Molecules. 2019;24(10):1892. PMID: 31100979. DOI: 10.3390/molecules24101892. PubMed
-
Dorr RT, Lines R, Levine N, Brooks C, Xiang L, Hruby VJ, Hadley ME. Evaluation of melanotan-II, a superpotent cyclic melanotropic peptide in a pilot phase-I clinical study. Life Sci. 1996;58(20):1777-84. PMID: 8637402. DOI: 10.1016/0024-3205(96)00160-9. PubMed
-
Mountjoy KG. Functions for pro-opiomelanocortin-derived peptides in obesity and diabetes. Biochem J. 2010;428(3):305-24. PMID: 20546242. DOI: 10.1042/BJ20091957. PubMed
Disclaimer. Statements in this article have not been evaluated by the Food and Drug Administration. This compound is not intended to diagnose, treat, cure, or prevent any disease. Sparta Labs sells research-use-only materials. Content is provided for educational and informational purposes only and does not constitute medical advice. Consult a qualified medical professional for any health concerns.
Frequently asked questions
Which melanocortin receptors does Melanotan-2 engage?
Published pharmacology characterizes MT-II as a non-selective agonist with reported activity at MC1R, MC3R, MC4R, and MC5R. It is not reported to meaningfully engage MC2R, the ACTH-specific receptor, because MC2R recognition depends on N-terminal ACTH residues that the truncated heptapeptide scaffold of MT-II lacks.
What is the significance of the lactam bridge in Melanotan-2?
MT-II carries an intramolecular lactam bridge that cyclizes the peptide and constrains the His-D-Phe-Arg-Trp pharmacophore into a rigid conformation. Structure-activity literature on constrained melanotropins associates this conformational restriction with high receptor affinity and resistance to enzymatic degradation relative to the linear alpha-MSH sequence.
What intracellular signaling pathway is associated with MT-II?
At each of its target subtypes, MT-II is reported to activate Gs-coupled signaling, stimulating adenylyl cyclase and raising intracellular cyclic AMP. Downstream, cAMP activates protein kinase A and, in pigment cells, CREB-dependent transcription. Reviews also note MC4R coupling to ERK1/2 MAP-kinase signaling as an area of continuing study.
Why does MT-II produce effects at both central and peripheral sites?
Melanocortin receptor subtypes differ in tissue distribution: MC1R is prominent in pigment cells while MC4R is concentrated in hypothalamic circuits. Because MT-II is non-selective, preclinical observations attributed to it span pigmentation biology and central energy-balance circuitry, and disentangling subtype- and site-specific contributions remains an active research task.