TB-500 Mechanism of Action: Published Research
A mechanism review of TB-500 (Ac-LKKTETQ) as a minimal fragment of thymosin beta-4: how the LKKTETQ WH2 module grips G-actin, why the activity needs no membrane receptor, and how actin buffering links to the cell-migration and NF-kB findings reported in peer-reviewed models. Educational reference.

For research use only. Not for human consumption. This article is educational reference material. It is not medical advice and is not a recommendation to use any substance.
From a 43-Residue Protein to a Seven-Residue Fragment
TB-500 is the synthetic, N-terminally acetylated heptapeptide Ac-LKKTETQ. Its sequence maps to residues 17–23 of thymosin beta-4 (Tβ4), a 43-amino-acid intracellular polypeptide that belongs to the beta-thymosin family of small, natively unstructured proteins. Because TB-500 is a deliberate excision of Tβ4's central segment rather than a novel molecule, its mechanistic literature cannot be read in isolation from that of the parent peptide.
The design logic is what makes this fragment interesting to structural biologists. Full-length Tβ4 folds around actin using three cooperating elements: an N-terminal helix, the central LKKTETQ segment, and a C-terminal extension. TB-500 isolates the middle element so that the contribution of that single actin-binding module can be studied apart from the flanking helices. This article reviews the reported molecular interactions and downstream observations for the fragment and its parent, with the level of evidence flagged in each case. A broader treatment of chemistry and regulatory context appears in the TB-500 research overview.
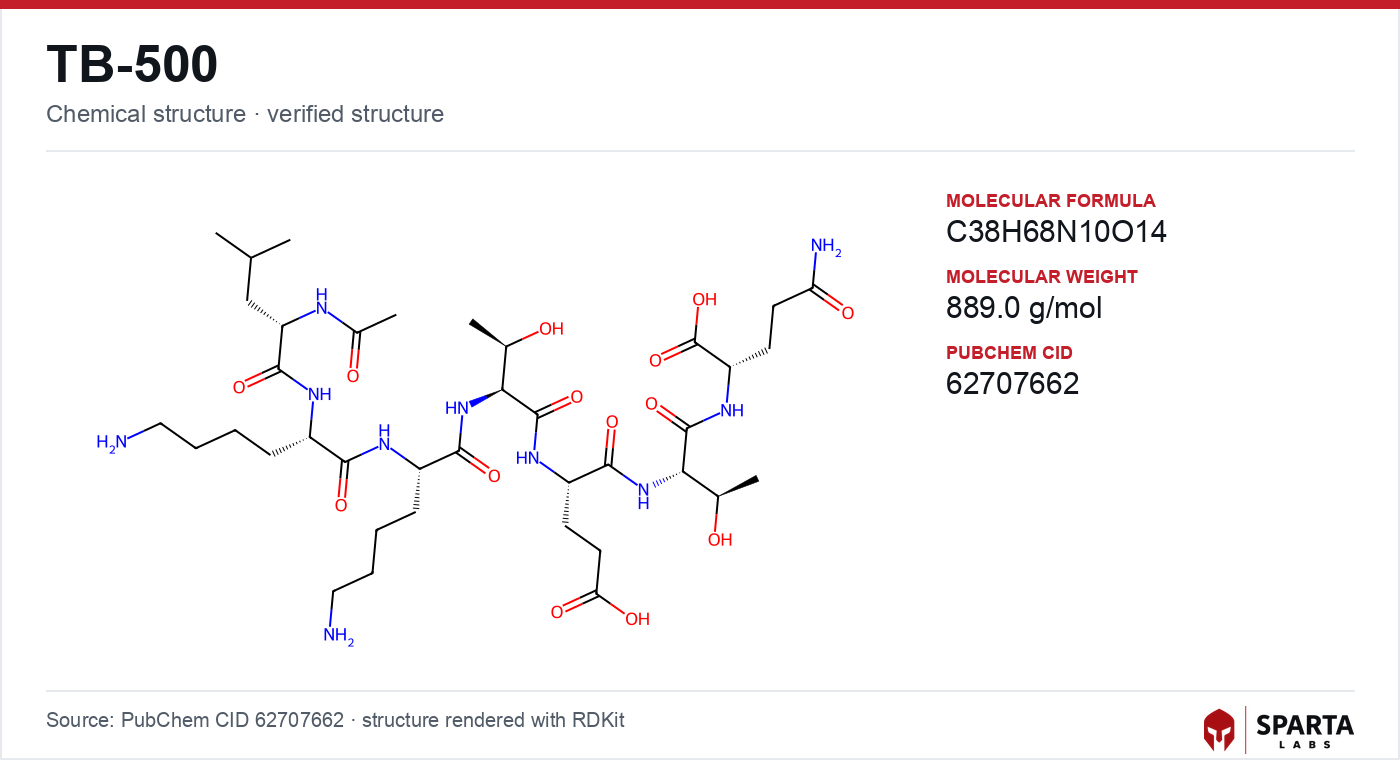
Figure: chemical structure of TB-500.
Actin Sequestration Without a Membrane Receptor
The defining feature of beta-thymosin pharmacology is that its best-characterized activity requires no classical membrane-bound receptor. Instead, the peptide acts inside the cytoplasm on the actin cytoskeleton itself.
The foundational biochemical work was published by Safer, Elzinga, and Nachmias in the Journal of Biological Chemistry in 1991, which reported that Tβ4 bound monomeric globular actin (G-actin) in a 1:1 molar ratio and inhibited its polymerization into filamentous actin (F-actin) in cell-free assays [1]. Complementary work reviewed by Safer and Nachmias established beta-thymosins as the principal intracellular G-actin buffer in several cell types, present at concentrations high enough to hold a substantial fraction of the monomeric actin pool in reserve [2].
Findings from research models do not establish safety or efficacy in humans. Sparta Labs makes no claims about the use of this compound.
The functional consequence described in this literature is a shift in the G-actin/F-actin equilibrium. By binding free monomers, the peptide reduces the pool available for filament elongation, a buffering role distinct from the severing and capping activities of other actin-regulatory proteins.
The WH2 Module: What LKKTETQ Actually Grips
A structural turning point came when the beta-thymosin actin-binding segment was recognized as a member of the WH2 (WASP-Homology 2) domain superfamily, a short motif shared with a large set of cytoskeletal regulators.
An X-ray crystallographic analysis published in the EMBO Journal in 2004 by Irobi and colleagues resolved how the beta-thymosin module contacts G-actin [3]. The study reported that the module engages the actin monomer across multiple surface patches at once, contacting both the barbed-end and pointed-end faces. This dual-contact architecture offered a physical explanation for why the peptide sequesters actin so effectively while preventing polymerization: it occludes the interfaces a monomer would otherwise use to join a growing filament.
Where the LKKTETQ core fits within that architecture was addressed directly by earlier truncation work. Safer and Chowrashi, publishing in Cell Motility and the Cytoskeleton in 1993, tested fragments of Tβ4 in the DNase I inhibition assay and reported that fragments retaining the LKKTETQ sequence held substantial G-actin-sequestering activity, whereas fragments lacking that central region showed markedly reduced activity [4]. That result is the mechanistic rationale for studying TB-500 as a minimal active unit rather than the full protein.
Reported Activities Beyond Actin Binding
Several molecular activities have been attributed to Tβ4 and, in some experiments, to the LKKTETQ fragment specifically. Each was observed in a defined model system, and the level of evidence varies.
Angiogenesis-associated activity
A study in the FASEB Journal in 2003 by Philp, Huff, Galli, Wei, Bhattacharya, Kleinman, and colleagues reported that the LKKTETQ sequence was associated with angiogenic activity in a chick chorioallantoic membrane (CAM) assay and in endothelial cell migration experiments in vitro [5]. The authors reported that the minimal fragment recapitulated a subset of the angiogenic activity attributed to full-length Tβ4 in the same preparations. These were embryo and cell-culture systems; the observations do not establish clinical outcomes.
NF-κB pathway modulation
A study in the Journal of the National Cancer Institute in 2003 by Cha and colleagues reported that Tβ4 interfered with tumor necrosis factor-alpha (TNF-α)-induced NF-κB activation in cell cultures, with evidence pointing to reduced nuclear translocation of the RelA/p65 subunit and altered downstream transcription [6]. These cell-model findings have not been confirmed in controlled human studies. A distinct inflammatory-signaling profile in cell models has been reported for GHK-Cu, another peptide studied within the healing and regenerative research cluster.
Epicardial progenitor mobilization
A study in Nature in 2007 by Smart, Risebro, Melville, Moses, Schwartz, Chien, and Riley reported that full-length Tβ4 stimulated outgrowth from quiescent adult murine epicardial explants, with differentiation into fibroblasts, smooth muscle cells, and endothelial cells [7]. This work used the intact 43-residue peptide; the relative contribution of the LKKTETQ fragment versus the flanking helices was not isolated in that publication, which is an important caveat when extrapolating to TB-500.
Corneal epithelial cell migration
Work reviewed by Sosne, Kleinman, and colleagues in Expert Opinion on Biological Therapy reported that full-length Tβ4 was associated with epithelial cell migration and wound closure in scratched corneal monolayer assays and rabbit corneal abrasion models [8]. Peptide-variant testing in that body of work identified LKKTETQ as the segment required for the migration activity.
How Actin Regulation Links to Cell Motility
The most mechanistically coherent thread across these reports is the connection between actin-monomer buffering and directional cell movement. A 2012 review in Expert Opinion on Biological Therapy by Goldstein, Hannappel, Sosne, and Kleinman described the linkage as follows: changes in the G-actin/F-actin equilibrium alter the leading-edge protrusion dynamics that drive cell migration, and Tβ4 modulates that equilibrium by controlling the pool of polymerizable monomers [9]. The authors characterized actin regulation as the most thoroughly established of the peptide's reported activities, with the other pathways at earlier stages of validation.
Framed this way, the reported effects on endothelial migration, corneal wound closure, and epicardial cell outgrowth are consistent with a common cytoskeletal node, though each remains an independent line of investigation. The parallels between actin-linked migration mechanisms across this compound class are examined further in the TB-500 published research summary.
Structural Limits of the Fragment and Open Questions
A recurring caveat in the primary literature is that TB-500 is not a full functional copy of its parent. Husson and colleagues, writing in the Annals of the New York Academy of Sciences in 2010, reviewed how the beta-thymosin/WH2 module operates within a larger structural context, noting that the N-terminal and C-terminal helices flanking LKKTETQ contribute independently to binding affinity and to the integrity of the peptide–actin complex [10]. TB-500 (Ac-LKKTETQ) lacks both flanking helices, so equivalence to full-length Tβ4 across every reported activity should not be assumed.
Several questions therefore remain open. The pharmacokinetic profile of the acetylated heptapeptide, including its metabolic fate, is not fully characterized. Many key observations rest on cell culture, rodent, or avian-embryo systems and await confirmation across additional biological contexts. And for activities first demonstrated with the intact protein, such as epicardial mobilization, the precise share attributable to the LKKTETQ fragment is unresolved.
Analytical specifications for research-grade Ac-LKKTETQ are documented on the product page for TB-500 from Sparta Labs, and verification-standard details are covered in the TB-500 sourcing and quality reference. Mechanistic comparisons with other peptides in the regenerative-research literature, such as the pathways discussed for BPC-157, provide additional context for how distinct molecular modes of action are described in the same field.
References
-
Safer D, Elzinga M, Nachmias VT. Thymosin beta 4 and Fx, an actin-sequestering peptide, are indistinguishable. J Biol Chem. 1991;266(7):4029–4032. PMID: 1999398. https://pubmed.ncbi.nlm.nih.gov/1999398/
-
Safer D, Nachmias VT. Beta thymosins as actin binding peptides. Bioessays. 1994;16(7):473–479. PMID: 8080474. https://pubmed.ncbi.nlm.nih.gov/8080474/
-
Irobi E, Aguda AH, Larsson M, Guerin C, Yin HL, Burtnick LD, et al. Structural basis of actin sequestration by thymosin-β4: implications for WH2 proteins. EMBO J. 2004;23(18):3599–3608. PMC517612. https://pmc.ncbi.nlm.nih.gov/articles/PMC517612/
-
Safer D, Chowrashi PK. Actin-sequestering ability of thymosin beta 4, thymosin beta 4 fragments, and thymosin beta 4-like peptides as assessed by the DNase I inhibition assay. Cell Motil Cytoskeleton. 1993;25(4):329–335. PMID: 8471179. https://pubmed.ncbi.nlm.nih.gov/8471179/
-
Philp D, Huff T, Galli U, Wei S, Bhattacharya B, Kleinman HK, et al. The actin binding site on thymosin beta4 promotes angiogenesis. FASEB J. 2003;17(14):2103–2105. PMID: 14500546. https://pubmed.ncbi.nlm.nih.gov/14500546/
-
Cha HJ, Jeong MJ, Kleinman HK. Role of thymosin β4 in tumor metastasis and angiogenesis. J Natl Cancer Inst. 2003;95(22):1674–1680. PMID: 14625257. https://pubmed.ncbi.nlm.nih.gov/14625257/
-
Smart N, Risebro CA, Melville AA, Moses K, Schwartz RJ, Chien KR, Riley PR. Thymosin beta4 induces adult epicardial progenitor mobilization and neovascularization. Nature. 2007;445(7124):177–182. PMID: 17108969. https://pubmed.ncbi.nlm.nih.gov/17108969/
-
Sosne G, Kleinman HK. Thymosin beta 4: a corneal wound healing and anti-inflammatory agent. Expert Opin Biol Ther. 2009;9(9):1107–1114. PMID: 19668473. https://pubmed.ncbi.nlm.nih.gov/19668473/
-
Goldstein AL, Hannappel E, Sosne G, Kleinman HK. Thymosin β4: a multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin Biol Ther. 2012;12(1):37–51. PMID: 22087795. https://pubmed.ncbi.nlm.nih.gov/22087795/
-
Husson C, Cantrelle FX, Roblin P, Didry D, Le KH, Perez J, et al. Multifunctionality of the β-thymosin/WH2 module: G-actin sequestration, actin filament growth, nucleation, and branching. Ann N Y Acad Sci. 2010;1194:44–52. PMID: 20536453. https://pubmed.ncbi.nlm.nih.gov/20536453/
Disclaimer. Statements in this article have not been evaluated by the Food and Drug Administration. This compound is not intended to diagnose, treat, cure, or prevent any disease. Sparta Labs sells research-use-only materials. Content is provided for educational and informational purposes only and does not constitute medical advice. Consult a qualified medical professional for any health concerns.
Frequently asked questions
How is TB-500 described at the molecular level?
In the published literature, TB-500 (Ac-LKKTETQ) is characterized as a G-actin sequestering peptide that binds monomeric globular actin in a 1:1 molar ratio and inhibits its polymerization to filamentous actin. This activity does not require a classical membrane-bound receptor and acts inside the cytoplasm on the actin cytoskeleton. The structural basis of the interaction has been resolved by X-ray crystallography.
Why is TB-500 studied as a fragment of thymosin beta-4?
TB-500 corresponds to residues 17 to 23 of the 43-amino-acid protein thymosin beta-4 and isolates its central LKKTETQ actin-binding module. Truncation studies reported that fragments containing LKKTETQ retained substantial actin-sequestering activity while fragments lacking it did not. This makes the fragment a way to study one binding module apart from the parent protein's flanking helices.
What is the WH2 domain and how does it relate to TB-500?
The actin-binding segment of thymosin beta-4 is a member of the WH2 (WASP-Homology 2) domain superfamily. Crystallographic work published in EMBO Journal in 2004 reported that this module contacts the actin monomer across multiple surface patches at once, engaging both the barbed-end and pointed-end faces. That dual-contact architecture is the reported explanation for its high-affinity, non-polymerizing sequestration of actin.
Do TB-500 findings from the parent peptide always apply to the fragment?
Not necessarily. Several activities, such as epicardial progenitor mobilization, were first reported using the full-length 43-residue thymosin beta-4 peptide, and the share attributable to the LKKTETQ fragment alone was not isolated in those studies. Reviews note that the N-terminal and C-terminal helices absent from TB-500 contribute independently to actin binding, so equivalence should not be assumed.